|
FROM FRANK MAGALLANES (Book review and OPINION are
my own). Readers are cautioned that they must read this online manuscript
(including the book above) in its entirety to form their own opinions:
Carl Popper (1966): As a realist I look
upon logic as the organon of criticisms (rather than of proof) in our
search for true and highly informative theories - or at least for new
theories that contain more information, and correspond better to the
facts, than our older theories. And I look upon criticism, in its turn,
as our main instrument in promoting the growth of our knowledge about the
world of facts. Source and references view.
Part of my responsibility at OPEFE is to review
the publication and evaluate any future problems for hobbyists who
collect these fish for the home aquarium. This review includes examining
the material and photographs to see if they correspond correctly to
imports of piranha fish. Also to evaluate the methods used to ascertain
whether or not the descriptions are correct and easy to understand. This
is very important for hobbyists and world wide fish dealers who rely on
OPEFE for that information in their day to day operations.
I was made aware of this species in 2005, however
I did not carry it on OPEFE until just recently. The following gives an
explanation; the material in its book form presents some interesting
views and potential problems with S. odyssei. The book uses
molecular and evolutionary perspectives. Beginning in the 1960s,
evolutionary biology was significantly transformed by the incorporation
of ideas and techniques from molecular biology. This led to many novel
views (and as many controversies) about phylogenetic relationships, rates
and mechanisms of evolutionary change, and standards of inference and
hypothesis testing VIEW.
The scientific literature from previous work was
included in the body of work by Hubert in the creation of this new
species. Much of the method is DNA related. The comparison is done
for this description with only 4 species; Serrasalmus
compressus (20 specimens) Bolivia, Madeira, Serrasalmus hastatus (11 specimens) Brazil,
Amazonas, Serrasalmus hollandi (24 specimens) Bolivia, Madeira,
and Serrasalmus altispinis (9
specimens) Brazil, Amazonas. What I don't understand is why S.
hollandi is even mentioned in this comparison. It is not a member of
the compressus group to my knowledge. Gery placed S. hollandi as a
member of the humeralis group in 1976. Then in 1977 revised it again as
follows; S. brandtii,
S. elongatus. S. hollandi, S. humeralis,
S. rhombeus,
S. rhombeus-marginatus,
S. sanchezi, S. spilopleura.
In the Hubert body of work, S. sanchezi has been moved to a
compressus group member, which I support based on the morphological
features. but because S. altuvei was not considered (see below) it raises
some taxonomic issues. S. geryi was not
included in the finished new description. It too is a member of the
compressus group.
Currently, two species remain difficult to
visually separate in the hobby, they are S.
maculatus and S. spilopleura.
These two species present morphology and locality issues since the
Jegu rehabilitation. The only helpful tool is to determine where the fish
was collected. Also problematic is S.
eigenmanni and S. humeralis.
While both those species are valid, there remains questions on the
taxonomy and especially the range. S. humeralis is found in Amazon
River basin: Bolivia, Brazil and Peru. S. eigenmanni is from the
Amazon River basin and northern and eastern Guiana Shield rivers: Brazil,
French Guiana, Guyana, Suriname and Venezuela. Both species are difficult
to differentiate and overlap their respective range. However, S.
eigenmanni is not found in Bolivia according to Eschmeyer (California
Academy of Sciences). Also problems with S. humeralis is the
species itself (more on this below), there is no Holotype specimen. I
have photos of this species by Adrian Leroy who stated Nicholas Hubert
identified the species for him (see S. humeralis). Yet these
species look nothing like the fishbase.org specimen photos. Not mentioned
by Hubert is Serrasalmus nigricans. (Spix & Agassiz 1829) Amazon River basin,
Brazil. This species is valid according Jegu in Reis et a., 2003:191 with
author as Agassiz. One wonders why this important species was overlooked
or not considered in the S. odyssei paper. Perhaps it was because
no types exist (Kottelat, 1988); See Kottelat, 988: 79 for details on
authorship. And because there is only a painted drawing of it and a
specimen with wrong label? VIEW
According to Hubert, S. spilopleura is
found in one river in Bolivia. However, in the Hubert map it overlaps S.
maculatus range; Amazon and Paraguay-Paraná River basin: Argentina,
Bolivia, Brazil, Colombia, Paraguay, Peru and Uruguay. Current
distribution for S. spilopleura is listed as; Guaporé River
basin, Paraná River basin: Argentina and Brazil according to Jegu, et
al.
Hubert further states that there is possible
hybridization between S. compressus and S. sp (= S.
odyssei). Hybridization between 2 sympatric species that don't spawn
together in polygyny is like our native sunfish (Lepomis), where
sperm drift is possible, would be rare indeed!?! One must wonder
about S. hollandi, especially since no one seems to be comfortable
with delineating what it really is.
Serrasalmus hollandi, according to CAS, is
a valid name. Its locality falls under questionable river's; Madeira
River basin and ? Guyana rivers: Brazil and Guiana (?).
S. hollandi has
not had any recent rehabilitation except in a few of lines of description
by Jegu in his rehabilitation of Serrasalmus maculatus. Photos
would be nice, but not seen in this book except for a preserved specimen
collected in 2005 (MNHN 2005-2268). The actual holotype of this species
is lost. I do not know if the preserved species is recognized as a new
holotype or simply a specimen that fits the Jegu/Hubert & Renno
description. Since it appears that S. hollandi is rarely
collected, begs the question, what is Hubert actually looking at? Is
it S. manueli Venezuela?
or this one Jegu S. humeralis?
or is this S.
humeralis in fishbase.org? Another S. manueli is from the
Rio Negro VIEW.
S. humeralis has historical description
problems such as contradicting descriptions and locality questions. One
can pick and choose which drawing or plate image to follow.
As one can see, both the Venezuela S. manueli
and the Amazonia S. humeralis at fishbase.org look strikingly
similar. If they are the same then S. manueli, Fernández-Yépez
& Ramírez, 1967 would likely be considered a synonym by some future
authority to S. humeralis Valenciennes in Cuvier et Valenciennes
1849 (Pl. XI, Fig. 2). Why? Because S. manueli is younger name
than S. humeralis, assuming both species were properly described.
This would make S. manueli a Venezuelan sister to S. humeralis Amazonia.
If they are not the same, then it would be up to a future Systematician
to diagnosis both species in a comparison study.
Hubert lists the cladogram (hypotheses) of
phylogenetic relationships with the characid subfamily Serrasalminae.
Part of the material for this species is derived from DNA analysis using
several species, including some questionable ones in the first body of
work. The author was unable to find any single diagnostic or
semi-diagnostic locus between S. eigenmanni and S. spilopleura.
He could not rule out that they might be different species. S.
eigenmanni is largely distributed in the upper Madeira basin,
but not in the upper Guapore River where only Serrasalmus spilopleura
has been found until now. Hubert et al 2006 showed the citation by
Jegu and Dos Santos 2001 from the Madeira was erroneous regarding S.
eigenmanni.
The error originated from confusion with S.
spilopleura (see Hubert & Renno 2009, 2010). Could this be S.
spilopleura specimen VIEW that Hubert said was
misidentified as S. eigenmanni by Jegu? This specimen, an unknown Pristobrycon-type
sp. collected by George Fear and photographed by David M. Schleser.
It has since been identified as S. serrulatus. It has a thin black
band and clear edge of the caudal fin. According Dave, this fish was
collected at the Tachshacurary river (a blackwater stream) just upstream
from the village of Tacshacuraray which is located where the
Tachsahcuraray empties into the Napo, Peru. David has caught many more in
all age classes.
Any advanced hobbyist who has seen S.
spilopleura and S. eigenmanni know those two species look
nothing like each other. But as a hobbyist, I personally wonder how Jegu
could have easily misidentify a specimen like S. eigenmanni and S.
spilopleura. But even S. eigenmanni has historical description
and locality problems (see further down below).
Ortega & Vari (1986) tentatively placed
specimens morphologically very similar S. spilopleura from
the Ucayali within S. humeralis, another closely related
species (Jegu & Dos Santos 2001).The results of the present
study by Hubert suggest that the specimens from the Ucayali analyzed here
currently belong to S. spilopleura. In my thoughts (whether right
or wrong), I suggest the image by Schleser fits the misidentified
specimen. Again, this is just a guess on my part and no scientific
license to say so. In dealing with bad scientific descriptions and
samples, one has to work with what one has. In this case, samples spread
out over the world that not just anyone can have access to. It would also
involve a team of researchers to explore all the world museums for
specimens and comparison's.
Also left to wonder why he mentions S.compressus
as ENDEMIC to Madeira...when it isn't an endemic species limited to that
river but wide spread species. Hubert, however offers an
explanation; through a combined use of biogeographic, phylogenetic and
phylogeographic approaches at several spatial scales (see back cover of
this book).
For hobbyists, biogeography is a relatively
new term in the field of science. It takes into account several
suppositions. It is usually considered to be part of physical geography
as it often relates to the examination of the physical environment
and how it effects species and shaped their distribution across space.
Below are applicable definitions;
DEFINITIONS
1. Biogeographic: (McGraw-Hill Science
& Technology Encyclopedia): The scientific study of the geographic
distribution of plant and animal life. Factors affecting distribution
include the geologic history of a region, its climate and soil
composition, and the presence or absence of natural barriers like
deserts, oceans, and mountains. Biotic factors such as interactions among
competing species, coevolutionary influences, and the reproductive and
nutritional requirements of populations and species are also
studied. A biogeographic region is a large, generally continuous
division of the Earth's surface having a distinctive biotic community.
Biogeographic regions are usually defined separately for floral and
faunal communities and are largely restricted to the terrestrial areas of
the Earth.
Comparative Biogeography
Discovering and Classifying Biogeographical
Patterns of a Dynamic Earth (2009)
Lynne R. Parenti, R. Claro and K.C. Lindeman
co-editor with M.L.J. Stiassny and G. D. Johnson.
(Quoted text) Description;
To unravel the complex shared history of the Earth and its life forms,
biogeographers analyze patterns of biodiversity, species distribution,
and geological history. So far, the field of biogeography has been
fragmented into divergent systematic and evolutionary approaches, with no
overarching or unifying research theme or method. In this text, Lynne
Parenti and Malte Ebach address this discord and outline comparative
tools to unify biogeography. Rooted in phylogenetic systematics, this
comparative biogeographic approach offers a comprehensive empirical
framework for discovering and deciphering the patterns and processes of
the distribution of life on Earth. The authors cover biogeography from
its fundamental ideas to the most effective ways to implement them.
Real-life examples illustrate concepts and problems, including the first
comparative biogeographical analysis of the Indo-West Pacific, an
introduction to biogeographical concepts rooted in the earth sciences,
and the integration of phylogeny, evolution and earth history.
Another good book to read : Biogeography and Plate
Tectonics (1987)
J.C. Briggs, Department of Marine Science,
University of South Florida, St. Petersburg, FL, USA
(Quoted text) Description;
One needs to look at only a small portion of the enormous literature on
plate tectonics published in the last 15 years to realize that there are
many differences between the various reconstructions that have been
presented. It becomes obvious that, although there is a general agreement
about the presence of an assembly of continents (a Pangaea) in the early
Mesozoic, there is considerable disagreement among earth scientists as to
the configurement of the assembly and the manner and timing of the
subsequent dispersal. While the revolution in geophysics was taking
place, systematic work in paleontology and neontology was being carried
out. This book is an attempt to incorporate the biological evidence into
the theory of plate tectonics. The author traces the changing
relationships among the various biogeographic regions and demonstrates
how such changes may often be correlated with the gradual geographic
alteration of the earth's surface. He analyses recent information about
the distribution of widespread groups of terrestrial and freshwater
vertebrates, invertebrates and plants, and discusses the biogeographical
effects of the movement of oceanic plates. It is particularly important
to obtain dependable information about certain critical times in the
history of continental relationships. We need to know when the
terrestrial parts of the earth were broken apart and when they were
joined together. The present investigation makes it clear that we cannot
depend entirely on evidence from plate tectonics nor will purely
biological evidence suffice. This book thus provides much of interest to
systematists working on contemporary groups of plants and animals,
paleontologists, evolutionary biologists, and professors teaching courses
in biogeography.
2. Phylogenetic: (Life Sciences &
Allied Applications / Biology) Biology the sequence of events
involved in the evolution of a species, genus, etc.
3. Phylogeographic: (Wikipedia English)
Phylogeography is the study of the historical processes that may be
responsible for the contemporary geographic distributions of individuals.
This is accomplished by considering the geographic distribution of
individuals in light of the patterns associated with a
gene genealogy.
So what exactly is Hubert telling us in his
book? For that you have to go back in time to Charles Darwin
and Alfred R. Wallace
One hundred and fifty years after the publication
of the Origin (Darwin), there is a revival of dispersal explanations,
occasioned in part by the development of molecular systematics and
molecular clocks. This is credited to Alfred Wallace work in the
mid-to-late 19th Century in England, as naturalist, explorer, geographer,
anthropologist, and biologist.. He extensively studied the Amazon River
and other land masses. He is today, considered the father of
biogeography.
(Quoted text) Molecular
dating of lineage divergence favours oceanic dispersal over tectonic
vicariance as an explanation for disjunct distributions in a wide variety
of taxa (de Queiroz, 2005). The molecular clocks mostly indicate that
divergences in phylogenetic trees occurred too recently to be explained
by vicariance. de Queiroz (2005) pointed out that although mismatches
between area cladograms and tectonic fragmentation do not necessarily
imply oceanic dispersal, in many cases dispersal is the most plausible
explanation. Thus in the past few years there has been an increase in the
number of molecular studies that support oceanic dispersal (e.g. Calsbeek
& Smith, 2003; Howarth et al., 2003; Heinicke et al., 2007; Vidal et
al., 2008). Therefore, dispersal hypotheses have claimed a new-found
respect, with molecular clock theory providing decisive evidence. But
there are those who strongly question the precise estimate of divergence
events by molecular clocks (e.g. Graur & Martin, 2004) and their
attributed evidence in favour of dispersal. (e.g. Heads, 2005b; Grehan,
2007; Nelson & Ladiges, 2009). view PDF
for conclusions and references.
As an additional foot note, biogeography and
the related sciences, raises red flags for believers in
creationism (Intelligent design) and a banner for atheists who do not
believe in creationism or God and support simple evolution of species
through processes. That is part of the controversy in researching this
field for hobbyists. I considered seriously whether or not to include
this in the page. But I asked myself, why not? It is part of the ongoing
debate.
Possible examples of specimens compared
to the new species S. odyssei
Below are samples of various caught fish
including their locality if available. Unfortunately, Hubert did not show
any live photos of the fish he references in his literature. Only 4
preserved specimens!
|

Serrasalmus compressus
Peru
Jégu [M.], Leão [E. L. M.] & Santos [G.
M. dos] 1991:102, Figs. 2 (a-b) [Ichthyological Exploration of
Freshwaters v. 2 (no. 2) Laguna Mocovi, Río Mocovi, Béni Province,
Bolivia. Holotype: MNHN 1986-0615;Paratypes: MNHN 1986-0616 (9, 1
c&s), 1986-0617 (3), 1986-0618 (1), 1986-0619 (1), 1988-1703 (3),
1988-1704 (1); INPA 1341 (3), 1342 (1), 1343 (1), 1344 (2), 1345 (1),
3327 (3); UTB (Univ. Técnica del Beni, Trinidad) uncat. (6), uncat.
(1). Valid as Serrasalmus compressus Jégu, Leão & Santos
1991 -- (Merckx et al. 2000:197, Hubert & Renno 2010:57. Current
status: Valid as Serrasalmus compressus Jégu, Leão &
Santos 1991. |
 \ \
Serrasalmus species resembling S. compressus but not as deep
bodied. Collected near Itionama river, Bolivia |

Serrasalmus compressus
Peru |

Serrasalmus maculatus,
San Martin river, Bolivia
Kner [R.] 1858:166 [6] [Sitzungsberichte der
Kaiserlichen Akademie der Wissenschaften.
Mathematisch-Naturwissenschaftliche Classe v. 32 (no. 22); Rio Guaporé,
Bobota, Mato Grosso, Brazil. Lectotype: NMW 57085. Paralectotypes: NMW
16344 (1, dry), 79457-59 (1, 1, 1, all dry). Appeared in more detail in
Kner 1860:43 [p. 35 of separate], Pl. 5 (fig. 11). Lectotype designated
by Jégu & Santos 2001:125. Valid as Serrasalmus spilopleura
Kner 1858 -- (Ortega & Vari 1986:9. Cestari & Galetti 1992,
Gómez & Chebez 1996:51, Britski et al. 1999:66, Butí &
Cancino 1999:71. Nakayama et al. 2000:149. Jégu & Santos 2001:119.
Lasso et al. 2001:96, Jégu in Reis et al. 2003:192. López et al.
2003:32. Casciotta et al. 2003:116. Menni 2004:78. Current status:
Valid as Serrasalmus spilopleura Kner 1858. |

Juvenile Serrasalmus sanchezi
Peru
Géry [J.] 1964:27, Fig. 22 [Beiträge zur
Neotropischen Fauna v. 4 (no. 1); "Caño Yarina", on edge of
Río Pacaya, tributary of Puinahua Canal, branch of lower Río Ucayali,
Peru. Holotype: ZFMK 1216. Paratypes: MHNG 2150.7 (1), ZFMK 1217 (1).
Type catalog: Busse 1984:217. Herder et al. 2010:118. Valid as Serrasalmus
sanchezi Géry 1964 -- (Ortega & Vari 1986:9. Jégu in Reis et
al. 2003:192. Current status: Valid as Serrasalmus sanchezi
Géry 1964. |

Adult Serrasalmus sanchezi (compare with
Hubert S. odyssei)
Peru |

Breeding Adult Serrasalmus sanchezi
Peru |
Serrasalmus humeralis types and historical
drawing. More information can be found by visiting the species page VIEW
With this species, it is unknown by me if a
Neotype has been erected. The Holotype is unique.
|
To see fishbase.org S. humeralis
Amazonia
VIEW
This one resembles S. manueli,
Venezuela
Valenciennes [A.] in Cuvier & Valenciennes
1850:279 [Histoire naturelle des poissons. v. 22; Amazonas, Brazil
[evidently in error for Araguay River, Tocantins basin]. Holotype
(unique): MNHN A-9735. Type catalog: Bertin 1948:25. See Jégu in Reis
et al. 2003:191 for type locality information. •Valid as Serrasalmus
humeralis Valenciennes 1850 -- Géry 1972:226, Ortega & Vari
1986:9, Géry et al. 1991:39, Planquette et al. 1996:376, Jégu
in Reis et al. 2003:191, Current status: Valid as Serrasalmus
humeralis Valenciennes 1850. Distribution: Amazon River basin:
Bolivia, Brazil and Peru. |

Text image Type de S. (Pristobrycon)
humeralis, 115 mm 1.sd. (dessin de H.-Ph. Danoy) Géry (1963)
Note black terminal band on caudal
Serrasalmus humeralis nob Another species
similar to that of M s in d Orbigny characterized by orbital in a
little closer they are striated and other parts opercular/ The lower
jaw is less prominent than the front of the well that it exceeds
the the entire width of the branch muzzle is rounded convex in front of
the nostril slightly concave eye audessus the interparietal crest is
convex,
1G B 4 d 16 A 33 P 7 V
color is blue for steel above the lateral line a
silver on the belly is marked black spot behind the hearing the back
and sides are covered with dark blue points almost blackish
caudal fin has a wide border Our type is black five-inch long it has
been reported from the Amazon by M Castelnau |

Serrasalmus species (humeralis?) from the Aruana (rio
Vermelho)
Note the V type caudal
This specimen appearance looks very much like a
humeralis type from Kner 1860. Except it does not have the
terminal caudal band. . |

Serrasalmus humeralis
Note the V type caudal
This is the Castelnau plate image of the
Holotype. It is mentioned in the IRD France data banks as part of the S.
humeralis specimen holotype collection.. Included is this
radiograph of the hototype MNHN A-9735: VIEW.
Likely used by Jegu, et al., depiction of S.
humeralis along with some specimens they believe to be the same in
the museum. According to the museum collection research I did, Jegu
moved a few specimens purported to be S. humeralis to S.
rhombeus.
As an advanced hobbyist, I'd be hard pressed to
say that plate image actually resembles anything of value. It's nicely
done. But with all the other descriptions and drawings, this is just
another one to add to the list of humeralis types. |

Serrasalmus humeralis
Zur Famiiie der Characine
III. Folge der Ichthyologischen Reitrage, Kner
1859 Fig. 9
See species page for full description.
Note the black terminal band on caudal
Serrasalmo humeralis.
Cuv. 8f Vol. xxii. p. 279; Kner, Denkschr. Acad. Wiss. Wien, 1860,
xviii. p. 38. taf. 4. fig. 9.
The height of the body is contained once and
five-sixths in the total length (without caudal), the length of the
head thrice. The second infraorbital is considerably longer than high,
and separated from the praeopercular limb by a strip of naked skin.
Abdominal serrature composed of from twenty-six
to twenty-eight spinous plates. Body with round blackish spots; a
large blackish blotch behind the gill-opening; caudal with the margin
black. |
Serrasalmus hollandi Holotype,
drawings and specimens
As I stated earlier, S. hollandi Holotype
is unique. I do not know if a Neotype has been erected for this species
yet. According to IRD databanks, M. Jegu has been determining S.
humeralis as Pristobrycon eigenmanni. Jegu determined 1
specimen as S. hollandi (MNHN 1989-1363) N. Hubert has
been collecting and determining his own specimens as S. hollandi.
|

Holotype Serrasalmus hollandi
Rio Guapore
Eigenmann [C. H.] 1915:251, Pl. 48 [Annals of
the Carnegie Museum v. 9 (nos. 3-4); Rio Guaporé at Maciél, Brazil.
Holotype (unique): FMNH 56978 [ex CM 5792]. Type catalog: Henn
1928:69, Ibarra & Stewart 1987:79, Valid as Serrasalmus
hollandi Eigenmann 1915 -- (Jégu & Santos 2001:136, Lasso et
al. 2001:96, Jégu in Reis et al. 2003:190, Machado-Allison et al.
2009:128, Hubert & Renno 2010:58, Current status:
Valid as Serrasalmus hollandi Eigenmann 1915. Distribution:
Madeira River basin and ? Guyana rivers: Brazil and Guiana (?). |

Serrasalmus hollandi - Eigenmann |

Juvenile species resembling S. hollandi
from El Prado, San Martin river, Bolivia
According to authority, this
may actually be juvenile of Serrasalmus spilopleura. |

Adult resembling S. hollandi El
Prado San Martin river, Bolivia .Maybe adult S. spilopleura. |

Juvenile S. hollandi? Pando, Bolivia |
Serrasalmus eigenmanni
Holotype is unique. Unknown by me if a
Neotype has been erected. Jegu & Santos, 2001 placed this species as
a Pristobrycon. However, Machado-Allison et al, 2009
returned it as a Serrasalmus. This species historically has been a
problem.
|
 \ \
S. eigenmanni Venezuela
Norman [J. R.] 1929:804, Fig. 16 [Proceedings
of the General Meetings for Scientific Business of the Zoological
Society of London 1928 (pt 4) (no. 30); Rockstone, Guyana. Holotype
(unique): BMNH 1911.10.31.496-497 (1 of 2, 120 mm). Norman mentions
only one specimen (type of the species) in the original description.
•Valid as Pristobrycon eigenmanni (Norman 1929) -- (Jégu
& Santos 2001:136, Valid as Serrasalmus eigenmanni
Norman 1929 -- (Géry 1972:224, Machado-Allison 2002:81,
Machado-Allison 2002:81, Machado-Allison et al. 2009:128, Current
status: Valid as Serrasalmus eigenmanni Norman 1929.
Distribution: Amazon River basin and northern and eastern Guiana
Shield rivers: Brazil, French Guiana, Guyana, Suriname and Venezuela. |

S. eigenmanni Venezuela |

Photo ID by M. Jégu
Messr. Jégu saw this photo and said the fish
was S. eigenmanni. W. L. Fink originally placed this as S.
serrulatus.
|
According to Hubert S. odyessi can be
confused with S. hollandi because both have the same body shape
and both reach the same standard length. He also states that S.
hollandi can be distinguished at least by the coloration pattern of
the caudal fin and the head, including meristic counts and morphometric
measurements. The S. hollandi caudal fin harbors a proximal
black band in young individuals up to 130 mm SL and this black band grows
in larger individuals, covering almost all the caudal fin. This
differentiation separates S. odyssei from S. hollandi with
the latter having just a terminal hyaline band in the distal part of the
caudal fin. Likewise, S. hollandi does not show the orange
coloration of the cheek nor does it have a higher number of scales
between the lateral line and the dorsal fin. Accordingly, S.
rhombeus is morphologically similar to S. odyssei and S.
hollandi., particularly for young specimens to 150 mm SL. But S.
rhombeus lacks the orange color of the cheek, presents a hyaline
caudal fin with a single terminal black band. And of course, specimens of
S. rhombeus have the red eye characteristic of the species. S.
compressus is closely related to S. hollandi and S.
odyssei, but is markedly distinct. S. compressus has a very
small head width, deeper body and lower number of prepelvic serrae.
|

Holotype
Serrasalmus serrulatus
Détail sur le(s) spécimen(s) : Nombre de
spécimen : 1 ;LS (mm) : 117 ;LT (mm) : 150 Conservateur : Alcool
Observations : HOLOTYPE DE PYGOPRISTIS SERRULATUS VALENCIENNES,1849
IN C.V.,H.N.P., XXII : 300 / VOIR REV. HYDROBIOL. TROP.,21(3) :
239-274,tabl. II Détail sur l'origine : Origine : Bresil Milieu :
Continent - Coordonnées : Bassin hydrologique : Amazone - Cours
d'eau : Amazone Provenance de la collection : Détail sur le(s)
collecteur(s) : Collecteur(s) : Castelnau ; Deville Date de
prélèvement : 1847 |
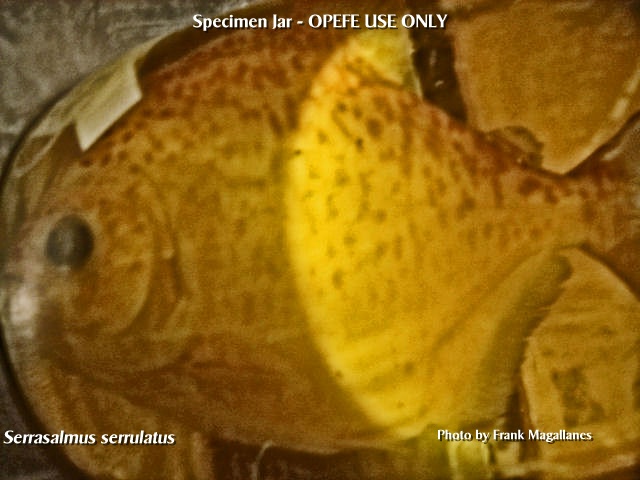
Serrasalmus serrulatus
UMMZ
I took this photo at the
University of Michigan during my visit with Dr. William L. Fink, 2006
Valenciennes [A.] in Cuvier &
Valenciennes 1850:300 [Histoire naturelle des poissons. v. 22;
Amazonas, Brazil. Holotype (unique): MNHN A-9898. Type catalog:
Bertin 1948:25-26, Valid as Serrasalmus serrulatus
(Valenciennes 1850) -- (Géry 1972:221, Ortega & Vari 1986:9,
Jégu in Reis et al. 2003:192, López et al. 2003:32, Menni 2004:78. Current
status: Valid as Serrasalmus serrulatus (Valenciennes
1850). Distribution: Amazon River basin and ? Essequibo River basin:
Amazon (?), Brazil, Guyana (?) and Peru; Argentina. |

Serrasalmus serrulatus
Collected rio Nanay, Peru
David M. Schleser
Locality data is on photo |

Serrasalmus serrulatus
Likely Peru
This is an aquarium specimen. |
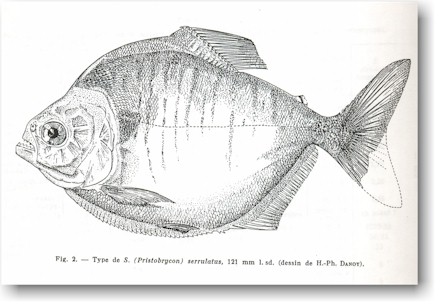
Serrasalmus serrulatus
This was a drawing in the Gery literature.
Information is on the image. |

Collected by Raúl Yalán, Rio Nanay, Peru
S. serrulatus. |
Hubert omitted S. altuvei because the
species is restricted to Venezuela so was not used for this body of work.
S. altuvei, S. hastatus, S. compressus, S. altispinis, and S.
geryi are members of the compressus group (Jegu et al.,). Hubert
revision; S. altuvei, S. hastatus, S. compressus, S. altispinis, S.
geryi, S. hollandi, S. sanchezi and S. odyssei.
|

Serrasalmus altuvei, Venezuela
Ramírez [M. V.] 1965:1, Fig. (p. 3)
[Evencias No. 14; El Polvero, Río San José, Guárico, Venezuela.
Holotype: MAC 65639. Paratypes: AFY 65307 (7); MAC 65627 (7);
MBUCV-V-12144 [ex MAC 65641] (1). Type catalog: Provenzano et al.
1998:17, Valid as Serrasalmus altuvei Ramírez 1965 -- (Jégu
et al. 1991:97, Taphorn 1992:328, Merckx et al. 2000:197, Fink
& Machado-Allison 2001:10, Machado-Allison 2002:68, Jégu in Reis
et al. 2003:190, Machado-Allison et al. 2009:128. Current status:
Valid as Serrasalmus altuvei Ramírez 1965.Distribution:
Orinoco River basin, Venezuela. |
The allopatric distribution patterns of
these two species and the absence of genetic differences between them
suggest that their taxonomy has been misunderstood (according to Hubert,
2010). Hubert does not compare S. sanchezi, also from the Ucayali
with this new species, reason unknown. Indeed, he mentions in passing
that S. sanchezi was split from the Ucayali and S. hastatus
from the Negro.
No further analysis was gathered to indicate why
he ignored S. sanchezi morphology in comparison to the new
species. Certainly, S. sanchezi is
comparable to S. odyssei in size and body shape. Though the colors
are distinctive, the opercle matches the deep coloration of both species.
Several other key features of S. sanchezi fit the new species
description. Hubert used DNA to state S. sanchezi and S.
compressus were related as sister.
I also find it most puzzling that
no comparison was done with S. sanchezi, a species found within
the range of S. odyssei and most similar in appearance to it.
Another species collected is Serrasalmus
iridopsis. Jacques Gery placed this as synonym of
S. humeralis. In my opinion, its
possible this may actually be a juvenile of either sanchezi or
odyssei.
|

Intenez River, 13 October 2002, 128
mm, Bolivia, by Nicolas HUBERT
Length: 16.7 cm SL male/unsexed
Hubert [N.] & Renno [J.-F.]
2010:49, 52 [Evolution of the Neotropical Ichthyofauna.; Rio Itenez,
Bella vista, 13°31.1'S, 61°33.2'W, Madeira, Bolivia. Holotype: MNHN
2005-2216. Paratypes: many at MNHN. •Valid as Serrasalmus odysei
Hubert & Renno 2010. Current status: Valid as Serrasalmus
odyssei Hubert & Renno 2010. Distribution: Rio San Martin and
Rio Paragá, Bolivia. |

Serrasalmus sanchezi HOLOTYPE
See credits on photo. |

Serrasalmus sanchezi, 144mm SL. Collected Amazon,
downstream from Iquitos, Peru |

Composite image of S. odyssei (top) Bolivia and Peruvian
Serrasalmus sp. collected as S. humeralis. The Peruvian
fish was collected with samples of S. sanchezi. Both are
sympatric. |
The book by Hubert documents specimens from
Bolivia in the Rio Madeira and other surrounding areas. The Hubert and
Renno used a German publication company that does not require peer review
according to Google search engines. A
method not often seen, but becoming more prevalent in a publish or perish
world. Why it was not published in a peer reviewed scientific
journal is unknown by me. Perhaps Hubert had his reasons. But at this
juncture, it doesn't matter.
The book is out there and can be purchased at
Amazon.com for those willing to spend the money for it. Overall, it is an
interesting book and gives this hobbyist a glimpse of the direction
piranha classifications is going. Ultimately, that is better than
nothing.
IN CONCLUSION
Added:
11/30/11
According to Historical Biogeography of
Neotropical Freshwater Fishes Edited by James S. Albert and Roberto E.
Reis 2011, Continental Analysis, page 161, they wrote: The suggestion
by Hubert and colleagues (2007a) that those intracratonic arches may have
played a role in determining vicariant events for the clade comprised by
Serrasalmus and Pygocentrus species should also be dismissed as a gross
misinterpretation of the nectectonic processes in the lower Amazon, which
were much more complex than a simple model of such deformation (e.g., J.
Crosta eat al., 2001). Though the lower Amazon River is a relatively
"recent" addition to the western-central Amazon River
ecosystem. Incorporated into the system since the breaching of the Purus
arch, dated either as taking place during the late Miocene. 8 MY
(Lundberg et al. 1998: Costa et al, 2001) or late Pliocene, -2.4 MY
(Campbell et al. 2006), faunistic differences between the portion of the
basin from the upstream reaches of the basin seems more likely to be due
to ecological rather than to historical, factors.
Until a future Systematician reviews the Hubert
species description, the species name is accepted and herein considered
valid.
REFERENCES
-
Hubert N. &
Renno J. F. 2010 Description of a new Serrasalmus species, Serrasalmus
odyssei n. sp. [pp. 52-59]. In: Evolution of the neotropical
ichthyofauna -- molecular and evolutionary perspectives about the
origin of the fish communities in the Amazon. VDM Publishing House,
Verlag Dr. Muller, Saarbucken, Germany. Evolution of the Neotropical
Ichthyofauna.: 95 pp.
-
See links above and OPEFE S. odyssei
-
Eschmeyer, W. N.
& Fricke, R. (eds.) Catalog of Fishes electronic version (30
November 2011). VIEW
-
Edited by James S. Albert and Roberto E. Reis
2011 Historical Biogeography of Neotropical Freshwater Fishes.
University of California Press. ONLINE.
RETURN USE
BACKSPACE OR CLICK HERE.
The OPEFE web site and its contents; is disclaimed
for purposes of Zoological Nomenclature in accordance with the
International Code of Zoological Nomenclature, Fourth Edition, Article
8.3 and 8.4. No new names or nomenclature changes are available from
statements at this web site.
Copyright© 1994-2012 Oregon Piranha Exotic Fish
Exhibit (The OPEFE fish exhibit is permanently CLOSED as of 2000)
Sutherlin, Oregon. Information posted on this web site is archival data
on fish scientific classifications and other information. DISCLAIMER: The
copyrighted material may not be used for any purpose other than private
study, scholarship or research. Cited information requires credit and
this link www.opefe.com. All rights
reserved. All images shown (unless otherwise noted) is property of
OPEFE. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}